quimico
2011-12-13 21:36
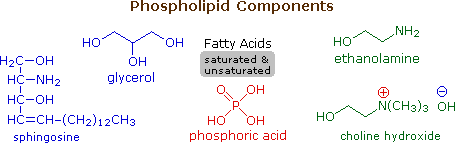
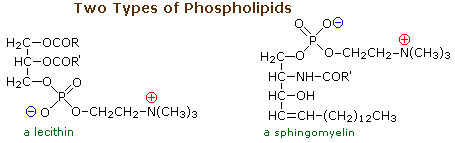
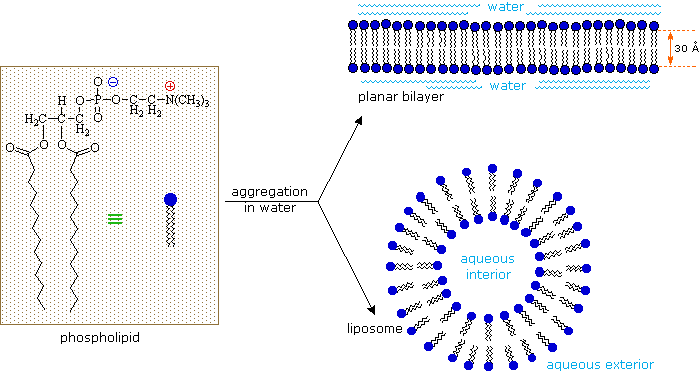
I lipidi sono una gruppo grande e diverso di composti organici ricorrenti in natura che sono correlati tramite la loro solubilità in solventi organici non polari (etere, cloroformio, acetone & benzene) e la generale insolubilità in acqua. C'è grande varietà strutturale tra i lipidi, come dimostrò prossimamente.
Acidi grassi
La caratteristica comune di questi lipidi è che sono tutti esteri di acidi grassi a catena da moderata a lunga. L'idrolisi acido o base catalizzata porta al componente acido grasso, alcuni esempi dei quali sono riportati sotto, assieme al componente alcole del lipide. Questi acidi carbossilici a lunga catena sono generalmente chiamati con nomi comuni, che nella maggior parte dei casi riflettono le loro origini. Gli acidi grassi naturali possono essere saturi o insaturi, e come indicato i seguenti dati, gli acidi saturi hanno m.p. più alti degli acidi insaturi di corripondente grandezza. I doppi legami nei composti insaturi elencati sotto sono tutti cis (o Z).
Saturi
CH3(CH2)10CO2H: acido laurico; m.p. 45 °C
CH3(CH2)12CO2H: acido miristico; m.p. 55 °C
CH3(CH2)14CO2H: acido palmitico; m.p. 63 °C
CH3(CH2)16CO2H: acido stearico; m.p. 69 °C
CH3(CH2)18CO2H: acido arachidico; m.p. 76 °C
Insaturi
CH3(CH2)5CH=CH(CH2)7CO2H: acido palmitoleico; m.p. 0 °C
CH3(CH2)7CH=CH(CH2)7CO2H : acido oleico; m.p. 13 °C
CH3(CH2)4CH=CHCH2CH=CH(CH2)7CO2H: acido linoleico; m.p. -5 °C
CH3CH2CH=CHCH2CH=CHCH2CH=CH(CH2)7CO2H: acido linolenico; m.p. -11 °C
CH3(CH2)4(CH=CHCH2)4(CH2)2CO2H: acido arachidonico; m.p. -49 °C
I punti di fusione più alti degli acidi grassi riflettono la forma uniforme tipo bacchetta delle loro molecole. Il doppio legame cis o i doppi legami cis negli acidi grassi insaturi introducono un nodo nella loro forma, che rende molto difficile impaccare le loro molecole assieme in una matrice stabile ripetitiva o nel reticolo cristallino. L'isomero con doppio legame trans dell'acido oleico, noto come acido elaidico, ha una forma lineare ed un m.p. di 45 °C (32 °C in più dell'isomero cis). Sotto sono mostrate le strutture degli acidi stearico ed oleico.
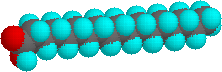
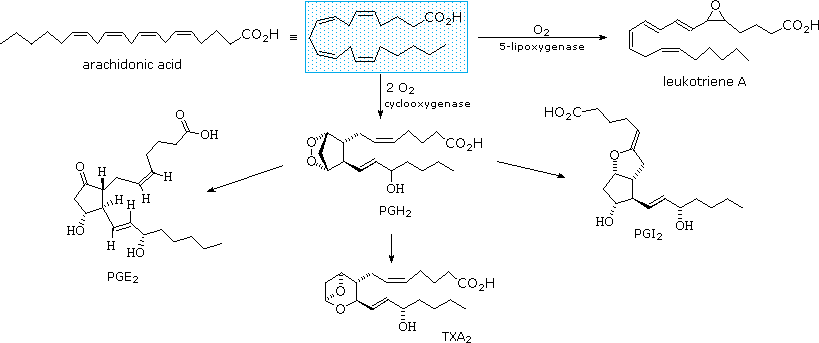
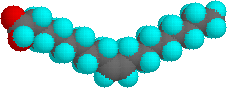 Due acidi grassi poliinsaturi, linoleico e linolenico, sono definiti "essenziali" perché la loro assenza nella dieta dell'uomo è stata associata a problemi di salute, come pelle squamosa, problemi di crescita e disidratazione eccessiva. Questi acidi sono anche precursori delle prostaglandine, una famiglia di lipidi fisiologicamente potenti presenti in quantità minime nella maggior parte dei tessuti del corpo.
A causa della loro aumentata acidità, gli acidi carbossilici reagiscono con basi a formare sali ionici, come mostrato nelle seguenti equazioni. Nel caso degli idrossidi dei metalli alcalini e di amine semplici (o ammoniaca) i sali risultanti hanno un pronunciato carattere ionico e sono di solito solubili in acqua. I metalli pesanti come argento, mercurio e piombo formano sali aventi maggiore carattere covalente, e la solubilità in acqua è ridotta, specialmente per acidi composti da quattro o più atomi di carbonio.
RCO2H + NaHCO3 → RCO2–Na+ + CO2 + H2O
RCO2H + (CH3)3N: → RCO2–(CH3)3NH+
RCO2H + AgOH → RCO2δ–Agδ+ + H2O
Due acidi grassi poliinsaturi, linoleico e linolenico, sono definiti "essenziali" perché la loro assenza nella dieta dell'uomo è stata associata a problemi di salute, come pelle squamosa, problemi di crescita e disidratazione eccessiva. Questi acidi sono anche precursori delle prostaglandine, una famiglia di lipidi fisiologicamente potenti presenti in quantità minime nella maggior parte dei tessuti del corpo.
A causa della loro aumentata acidità, gli acidi carbossilici reagiscono con basi a formare sali ionici, come mostrato nelle seguenti equazioni. Nel caso degli idrossidi dei metalli alcalini e di amine semplici (o ammoniaca) i sali risultanti hanno un pronunciato carattere ionico e sono di solito solubili in acqua. I metalli pesanti come argento, mercurio e piombo formano sali aventi maggiore carattere covalente, e la solubilità in acqua è ridotta, specialmente per acidi composti da quattro o più atomi di carbonio.
RCO2H + NaHCO3 → RCO2–Na+ + CO2 + H2O
RCO2H + (CH3)3N: → RCO2–(CH3)3NH+
RCO2H + AgOH → RCO2δ–Agδ+ + H2O
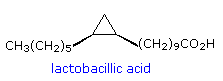
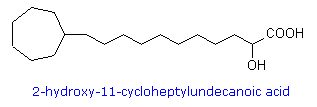
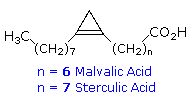
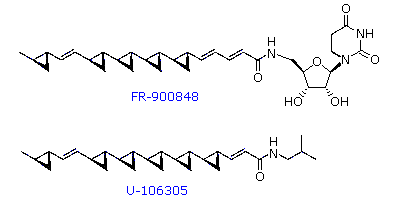
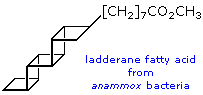
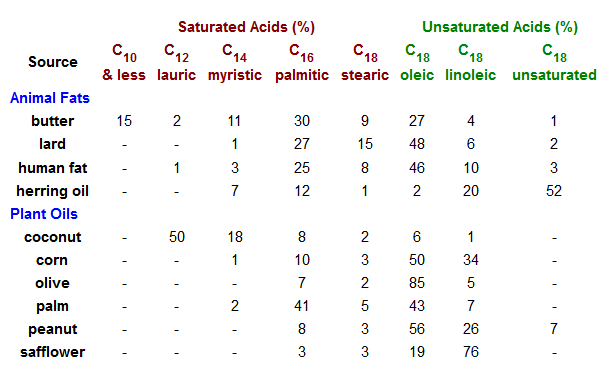

 possono essere sature o insature.
possono essere sature o insature.