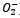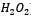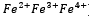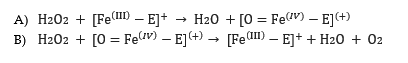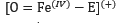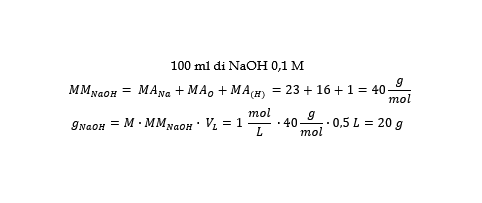Myttex Forum ha chiuso definitivamente. Non è più possibile inviare messaggi, ma il contenuto è ancora consultabile in questo archivio.
Mercaptano
2019-12-31 17:22
L'attività enzimatica del fegato, in qualunque essere vivente che lo possegga, è di una complessità enorme, e pertanto non mi propongo di trattarla tutta in maniera esaustiva, ma per quel che riguarda un enzima in particolare, ovvero la catalasi, ho avuto modo di effettuare vari saggi su campioni biologici freschi (Bos Taurus).
Di seguito, la relazione che ne ho tratto:
25/09/2019
Obiettivo: Studiare il comportamento dell’enzima catalasi.Verificare la presenza della catalasi in un campione di fegato bovino.Studiare la sua funzione e se essa si modifica con l’intervento di calore e variazione di pH.
Materiali e attrezzature:
-Provette da batteriologia.
-Portaprovette.
-Bisturi.
-Becher.
-Matraccio da 500 ml.
-Bagnomaria.
-Spatola.
-Bacchetta di vetro.
-Vetrino a orologio.
-Bilancia digitale (S: 0,01 g P:500 g).
-Fegato di bovino.
-HCl al 37% v/v (corrosivo, tossico per inalazione)
-NaOH in pastiglie (corrosivo, tossico)-Blu di metilene (Nocivo, irritante).Principio del metodo: Gli enzimi sono biomolecole con struttura proteica che ricoprono la funzione di biocatalizzatori nei confronti delle reazioni che avvengono all’interno di tutti gli organismi viventi.Dal punto di vista energetico, agiscono diminuendo l’energia di attivazione necessaria all’inizio del processo chimico che devono catalizzare, interagendo in maniera molto specifica con i reagenti che vi prendono parte secondo un modello chiave-serratura o un modello a adattamento indotto.Essenzialmente, gli enzimi agiscono abbassando l’energia libera degli stadi di transizione (ΔG‡), stabilizzando la struttura degli intermedi di reazione.Dal punto di vista chimico, si tratta di proteine che, grazie ai loro siti attivi e alle loro caratteristiche strutturali, riescono a catalizzare reazioni che altrimenti necessiterebbero di condizioni incompatibili con la vita (alte temperature, alte pressioni, impiego di corrente elettrica ad alto voltaggio).catalaseee.PNG
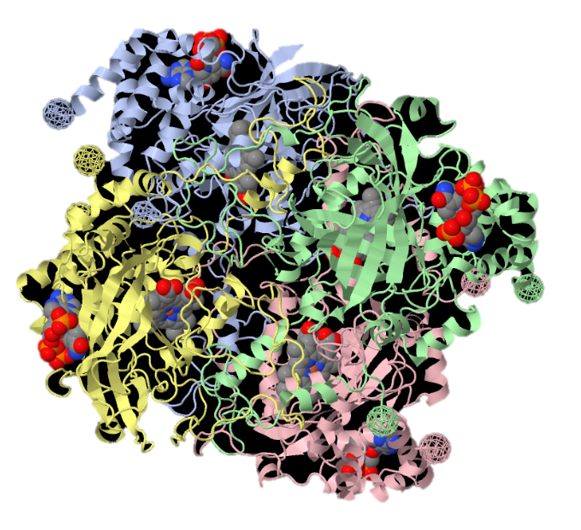
Figura 1, Catalasi bovina 4blc, modello tridimensionale da cristallografia a raggi X. Da proteopedia, enciclopedia digitale delle strutture proteiche.
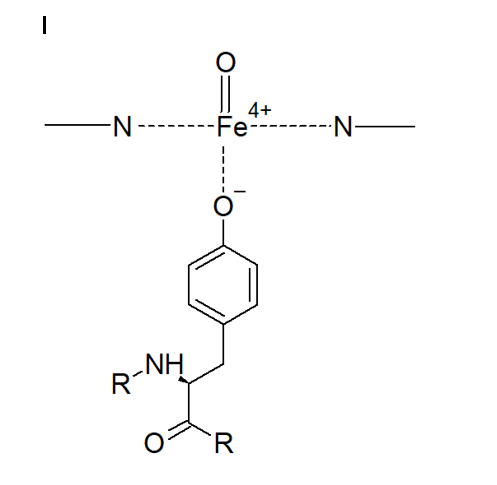
L’enzima catalasi, o per meglio dire, la famiglia di isoforme degli enzimi catalasi, appartengono alla classe delle ossidoreduttasi (EC 1), in particolare alla sottoclasse delle perossidasi (EC 1.11.1), il carattere particolare dell’enzima (l’azione di dismutazione sul perossido di idrogeno) determina la sua classificazione come EC 1.11.1.6. Insieme alla superossido-dismutasi, la catalasi agisce come agente antiossidante nei confronti delle specie reattive dell’ossigeno (ROS), le quali si vengono spesso a generare sia per intervento di agenti chimici o fisici esterni alla cellula, sia come scarto di alcuni processi biochimici.Nello specifico, la superossido-dismutasi agisce nei confronti dei radicali liberi superossido (anioni superossido superossido.PNG) catalizzando per la loro dismutazione in perossido di idrogeno (peross.PNG) e ossigeno molecolare (O2).La catalasi, invece, agisce sul primo prodotto della reazione precedente, catalizzando la sua dismutazione in acqua e ossigeno molecolare.Dal punto di vista strutturale, la catalasi è una emoproteina quaternaria; più specificatamente, un tetramero presentante quattro subunità identiche intrecciate tra loro.Tale conformazione garantisce la stabilità dei complessi enzimatici nel corso dei meccanismi di reazione (i cui reagenti e intermedi sono particolarmente energetici).Ognuna di queste subunità è legata a un gruppo eme B (Fe-protoporfirina IX), al cui interno si trova uno ione ferro, il cui numero d’ossidazione (+2, +3 o + 4, presenti rispettivamente infe.PNG) varia in funzione della forma assunta dall’enzima.Le differenze strutturali tra le forme ossidate e ridotte del substrato enzimatico svolgono un ruolo chiave nel processo di dismutazione.Nell’eme B della catalasi bovina (quella impiegata in questa esperienza), il ferro è coordinato sul piano dell’anello protoporfirinico dai quattro atomi d’azoto delle subunità pirroliche (prime 4 posizioni di coordinazione), ma presenta un totale di 6 legami di coordinazione, in quanto la quinta e la sesta posizione, sono occupate rispettivamente dal fenolato di un residuo di Tyr e dall’acqua. A differenza che nell’emoglobina, nella catalasi, il ferro è stato accertato trovarsi coordinato in forma stabile e non nociva come Fe3+ (presente comunque anche in forma di Fe2+), ed è attualmente accettata la teoria che prevede l’impiego del gruppo eme con ione ferrico coordinato come effettivo catalizzatore della reazione. La teoria più accreditata, difatti, si basa sulla presenza di altre 3 diverse forme strutturali enzimatiche (composti I, II e III) che differiscono dalle due strutture canoniche esclusivamente per lo stato d’ossidazione del ferro contenuto nell’eme e dei gruppi ad esso legati. Tali composti non sono altro che intermedi della reazione di dismutazione, utili a spiegare anche il meccanismo della stessa.È accertato lo svolgersi del processo in due diversi passaggi chiave, uno di ossidazione e uno di riduzione.Inizialmente il perossido d’idrogeno interagisce con l’Fe3+ dell’eme, portando alla produzione di acqua per riduzione del perossido e del Composto I per ossidazione dell’eme, ovvero [O=Fe4+(P-)].coordinazione catal.PNG
Figura 2, Probabile struttura del primo intermedio di reazione (composto I) coordinato dagli atomi d’azoto porfirinici e dal fenolato del residuo Tyr.
R rappresenta il proseguire della catena polipeptidica
Si tratta di un complesso nel quale il ferro presenta un doppio legame altamente instabile con un ossigeno caricato positivamente, la struttura si ritiene essere stabilizzata grazie alla presenza del ligando fenolato (residuo di Tyr), in grado di bilanciare la struttura grazie alla sua carica negativa sull’ossigeno coordinante.Nel secondo passaggio il Composto I interagisce con una seconda molecola di perossido, la quale agisce stavolta da riducente, portando il ferro a N.O. +3, e ripristinando in tal modo la struttura originaria del catalizzatore (enzima) e portando a compimento la reazione, con la liberazione di O2 e H2O.È stato supposto che la istidina e l’asparagina (o l’aspartato), presenti nel sit o attivo possano agevolare il distacco della molecola di O2 nel corso del secondo passaggio, oltre che lo spostamento di un protone del perossido d’idrogeno nella posizione intermedia di legame tra i due ossigeni (primo passaggio), in modo da favorire la formazione di una molecola d’acqua e del legame tra l’ossigeno deprotonato e il ferro dell’eme.Il composto I è di colore verde, e si presenta come una forma ossidata dell’enzima ferrico, la sua presenza come intermedio di reazione è molto veloce, abbastanza da non essere identificabile ad occhio nudo durante il maccanismo di reazione.La sintesi del gruppo eme contenuto nella catalasi avviene nei mitocondri ed è fondamentale la sua presenza all’interno di quasi tutte le specie viventi aerobie, proprio perché ricopre un ruolo fondamentale come componente dei siti attivi della catalasi (anch’essa ubiquitaria in quasi ogni specie aerobia).In particolare, si trovano alte concentrazioni di questo enzima nelle strutture biologiche dei mammiferi che sintetizzano o necessitano alto apporto di eme (fegato e globuli rossi). Per tale motivo si eseguono test sulle caratteristiche della catalasi soprattutto su porzioni di fegato.Referenze (sitografia e bibliografia):(VEDI ULTIMA PAGINA)
Procedimento operativo:
Il procedimento richiede l’utilizzo di idrossido di sodio, per alcuni dei saggi sulla reattività dell’enzima in ambiente alcalino.
Per tale motivo, se ne prepara una soluzione 1 M da 500 ml:
a) Si prelevano 20 g di NaOH solido dal suo contenitore mediante bilancia digitale.
b) Si discioglie il solido in un volume di acqua inferiore a quello prefissato per la soluzione.
c) Si mescola con una bacchetta di vetro pulita fino alla completa dissoluzione del soluto.
d) Si attende che la temperatura cali (la dissoluzione è esotermica).
e) Si travasa in un matraccio da 500 ml e si porta a volume con H2O distillata.
f) Si tappa il matraccio, si capovolge per omogenizzare la soluzione e si riappoggia sul bancone.
g) Infine, si scrivono le informazioni riguardanti contenuto, operatore e data di preparazione della soluzione sul vetro con un pennarello.
Preparata la soluzione, si procede con i preparativi dell’esperienza effettiva:
a) Si preleva una porzione più piccola di fegato dall’organo intero.
b) Si pone tale porzione su un foglio di plastica (in modo da non sporcare il bancone).
c) A partire dalla porzione prelevata, si sezionano in maniera fine delle strisce di tessuto epatico, in modo da ottenerne 7 porzioni di dimensioni simili.
L’esperienza si divide in 2 parti distinte:
a) Analisi in ambiente neutro.
b) Analisi in ambienti acido e alcalino.
Analisi in ambiente neutro.
a) Si preparano 6 provette e le si pone in un porta-provette.
a. Nelle prime 2 si pone solo il fegato.
b. Nelle successive 2 fegato e una goccia d’indicatore (blu di metilene).
c. Nelle ultime 2 si versa H2O distillata per 1/3 del volume totale. (Queste ultime saranno impiegate per testare la purezza dell’H2O2.)
b) Si dividono quindi le 6 provette in due gruppi da 3. Nelle prime tre si lavorerà a freddo. Nelle altre 3 si lavorerà a caldo (tra i 90 e i 100 °C) e per tale motivo le si pone in termostato mentre si lavora sulle altre.
c) Il procedimento, sia a caldo che a freddo, è lo stesso:
d) Si pongono alcuni ml di H2O2 nella provetta (sia che contenga fegato sia che contenga H2O) e si annotano le relative osservazioni.
Analisi in ambienti acido e alcalino.
a) Si lavano 2 delle provette precedentemente impiegate e vi si pongono 2 delle restanti 3 porzioni di fegato. Quindi si procede in maniera differente per ognuna delle 2.
i. Nella prima provetta si aggiunge 1 ml di HCl al 37% v/v mediante una pipetta e lavorando sotto cappa, data l’emissione di vapori corrosivi dal contenitore.
ii. Nella seconda provetta si aggiunge NaOH 1M fino a riempire 1/3 della provetta.
b) Si procede ad aggiungere in ognuna delle 2 provette una quantità uguale di H2O2 e si annotano le relative osservazioni.
c) Si ripete la prova in ambiente alcalino utilizzando l’ultima porzione di fegato rimasta, con la stessa procedura indicata sopra, ma lavorando a caldo.
d) Si annotano le osservazioni relative anche a questa prova.
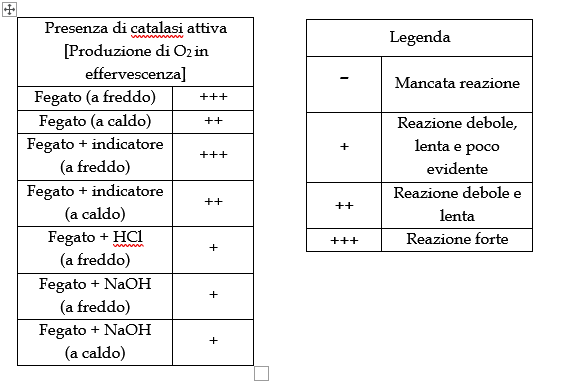
La reazione che dovrebbe intercorrere è la seguente (già bilanciata).eq. catalas.PNGCon le seguenti semireazioni (N.O. evidenziato in blu per non confonderlo coi coefficienti stechiometrici):eq catal2.PNGNel caso specifico della reazione catalizzata da enzima catalasi, l’equazione di reazione più accreditata coinvolge il gruppo eme, in quanto il meccanismo decorre in due passaggi:eq catal3.PNGDove abac.PNG è una rappresentazione semplificata del gruppo eme abachio.PNGè una rappresentazione semplificata di un composto di coordinazione intermedio con ferro tetravalente (a sua volta forma mesomerica di Fe(V)-E creatasi grazie alla parziale capacità di dispersione della carica nel sistema aromatico del gruppo eme).Calcoli e tabelle:tabella1.PNGNAOH.PNGImmagini:IMMAg.PNGOsservazioni: A temperatura ambiente e allo stato solido, l’idrossido di sodio si presenta sotto forma di pasticche bianche e opache. Una volta in soluzione, libera una discreta quantità di energia sotto forma di calore (dissoluzione esotermica). La soluzione ottenuta si presenta inodore, incolore e trasparente.L’HCl al 37 % v/v si presenta come un liquido volatile, inodore, incolore e trasparente. Dal contenitore si emana costantemente un aerosol corrosivo e per tale motivo, lo si impiega prettamente sotto cappa. Il blu di metilene (qui impiegato in soluzione concentrata con etanolo) è un composto dall’intenso colore blu.Il fegato impiegato, si presenta di colore bordeaux abbastanza intenso, con una netta differenza tra l’intensità e la brillantezza del colore esterno rispetto a quello interno.L’esterno è di fatto non solo più esposto ai processi di deterioramento dei materiali biologici, ma è anche rivestito da una membrana connettivale detta capsula di Glissman.Il perossido di idrogeno al 3% si presenta come un liquido inodore, incolore e trasparente.Per quanto riguarda l’esperienza vera e propria:a)Aggiungendo qualche ml di perossido al solo fegato a temperatura ambiente si ottiene una vigorosa reazione di dismutazione, nella quale il composto di partenza si disproporziona in acqua e ossigeno. La reazione è abbastanza forte da produrre un’effervescenza tale da far quasi traboccare le bollicine dal bordo della provetta.b)Aggiungendo qualche ml di perossido al fegato riscaldato (prova a caldo), si ottiene lo stesso effetto, ma con un vigore e una velocità di reazione decisamente inferiori. Da ciò si deduce che quantitativamente una parte dell’enzima si sia denaturata (minor vigore della reazione), mentre la restante parte si trovi al di fuori del suo optimum, catalizzando in maniera poco efficiente la reazione (velocità di reazione rallentata).c)Ripetendo la stessa prova in presenza di metiltioninio cloruro (blu di metilene) si ottengono i medesimi risultati, senza che però si presenti alcun tipo di viraggio da blu a incolore, indicando il mantenimento di un ambiente redox ossidante (sia a caldo che a freddo).d)Aggiungendo qualche ml di perossido all’H2O distillata (sia a caldo che a freddo), non si apprezza alcun cambiamento fisico, al di là dell’aumento di volume del liquido nella provetta, ciò indica che l’acqua ossigenata non sia contaminata chimicamente o peggio, già disproporzionata. e)Aggiungendo a freddo 1 ml di HCl al 37% v/v alla provetta contenente fegato, avviene un immediato cambiamento di colore dal trasparente al bianco semi-opaco, con la formazione di una sospensione di precipitati diversi, tutti di colore chiaro, probabilmente proteine denaturate dall’azione dell’acido.Il fegato contenuto nella provetta si presenta come decolorato, dal colore biancastro. Se ne deduce che tutte le sostanze che vi donavano colore risultano degradate a pH di 1 o inferiori (misura effettuata mediante cartina al tornasole, colorazione rosso-bruna).Aggiungendo qualche ml di perossido di idrogeno alla provetta si ottiene una reazione a malapena visibile, con formazione di pochissime bolle di O2.f)Aggiungendo NaOH 1M (per circa 1/3 del volume) a una provetta contenente fegato, non si apprezzano significativi cambiamenti nell’apparenza del campione.Aggiungendovi qualche ml di perossido, si può notare la leggerissima formazione di una effervescenza di O2, segno che nemmeno un pH pari a 13 o più sia in grado di denaturare del tutto l’enzima catalasi.g)Ripetendo la precedente prova a caldo, si ottengono risultati simili, con una reattività ancora minore (formazione di pochissime, circa 4, bolle di O2).Conclusioni: In base a quanto osservato, si può dedurre che la catalasi è un enzima insolitamente stabile ad alte temperature e sbalzi di pH anche notevoli. In ambiente alcalino, non denatura del tutto, portando alla formazione di qualche bolla di O2e non si denatura del tutto nemmeno a pH di 1 e inferiori, dismutando parte del perossido di idrogeno.Risulta particolarmente resistente alle alte temperature rispetto al resto delle proteine, con una temperatura di denaturazione che risulta superiore ai 90 °C (come si evince dalle prove eseguite a caldo).Riguardo al saggio sull’ambiente redox in cui avviene l’azione della catalasi (aggiunta di indicatore), data la mancanza di alcun tipo di viraggio (forse a causa del metodo di catalisi) si può assumere che esso resti sempre ossidante.Questo anche perché si ricorda che il sistema chimico di riferimento, contiene comunque un esigua quantità di perossido e che la dismutazione di quest’ultimo porta all’allontanamento di uno dei due prodotti in quanto gassoso, lasciando in soluzione soltanto acqua, proteine e catalasi. Referenze (bibliografia e sitografia):a)https://proteopedia.org/wiki/index.php/Catalase#cite_ref-4b)↑ Melik-Adamyan W, Bravo J, Carpena X, Switala J, Mate MJ, Fita I, Loewen PC. Substrate flow in catalases deduced from the crystal structures of active site variants of HPII from Escherichia coli. Proteins. 2001 Aug 15;44(3):270-81. PMID:http://www.ncbi.nlm.nih.gov/pubmed/1145560011455600c)Bertini, Ivano and Gray, Harry B. and Lippard, Stephen J. and Valentine, Joan Selverstone (1994) Bioinorganic Chemistry.University Science Books , Mill Valley, CA. ISBN 0-935702-57-d)https://chem.libretexts.org/Bookshelves/Inorganic_Chemistry/Book%3A_Bioinorganic_Chemistry_(Bertini_et_al.)/5%3A_Dioxygen_Reactions/Catalase_and_Peroxidasee). http://resolver.caltech.edu/CaltechBOOK:1994.002f)https://oca.weizmann.ac.il/oca-docs/fgij/fg.htm?mol=4BLC
I seguenti utenti ringraziano Mercaptano per questo messaggio: LuiCap, zodd01, TrevizeGolanCz, Claudio, luigi_67, A.Berenini, ClaudioG., Beefcotto87
luigi_67
2020-01-07 09:29
Oltre al semplice "grazie" volevo ringraziarti "per estreso" per aver condiviso questa interessante esperienza, descritta tra l'altro anche molto bene... l'ho letta un po' di corsa ma mi riprometto di ripassarmela con più calma.
Continua pure così !!!
Un saluto
Luigi
I seguenti utenti ringraziano luigi_67 per questo messaggio: Mercaptano
Oltre al semplice "grazie" volevo ringraziarti "per estreso" per aver condiviso questa interessante esperienza, descritta tra l'altro anche molto bene... l'ho letta un po' di corsa ma mi riprometto di ripassarmela con più calma.
Continua pure così !!!
Un saluto
Luigi
Grazie mille! Spero di poter aggiungere più discussioni simili a questa e inerenti biologia e biochimica, il laboratorio scolastico si sta attrezzando proprio in questi mesi per condurre nuove esperienze.
OT: Conto di postare qualcosa sulla PCR entro metà del prossimo quadrimestre scolastico.